


Seneste nyheder

klima
17.04.2024
Verdens tundra udleder overraskende mere CO2, når det bliver varmere

Mental sundhed
17.04.2024
Svingende kaffepriser presser vietnamesiske bønder mentalt

Kvantedata
15.04.2024
Internettet kan få kvantefart med lys gemt som lyd

ARTER
12.04.2024
Hegn og veje har givet ikonisk savannedyr genetiske problemer

Sonningprisen
11.04.2024
Efter blodprop: Marina Abramović får nu overrakt Sonningprisen

udvikling
11.04.2024
Økonom: Vi overser varer fra skoven for milliarder af kroner
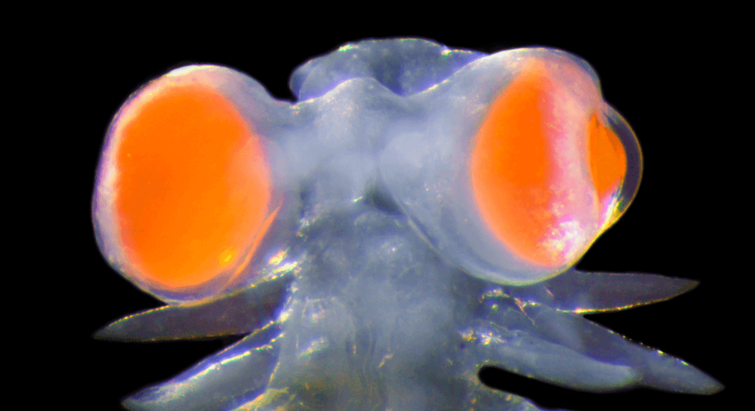
Havbiologi
09.04.2024
Middelhavs-orm har udviklet forunderlige øjne ”så store som møllehjul”

Landbrug
05.04.2024
”Drop industri-landbrug”: Kæmpestudie fastslår at mennesker og miljø har bedst af varieret landbrug, men bundlinjen trives også
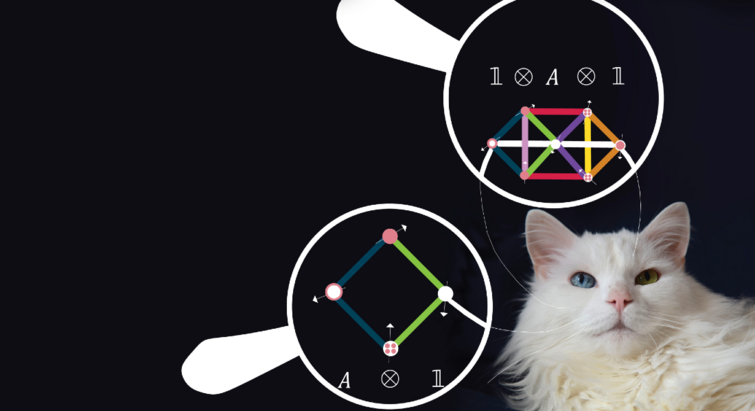
Kvantepartikler
04.04.2024
”Det handler i sidste ende om at kunne forudsige alt” – teori kan vise vej til forjættede kvantematerialer

kunstig intelligens
03.04.2024
Forskere viser vejen: AI-modeller behøver ikke at sluge SÅ meget strøm

Grøn data
26.03.2024
Det er klima-dyrt at opbevare sundhedsdata. Så i fremtiden skal vi måske kode det ind i DNA

Grøn medicin
26.03.2024